
编者按:年龄相关性黄斑变性(AMD)是老年人视力障碍的主要原因,目前眼科医师对其发病机制了解有限,所以缺乏有效的治疗方法。AMD的进展最初以视网膜色素上皮(RPE)的萎缩性改变以及溶酶体脂褐素和细胞外核黄素沉积形成为特征,慢性氧化应激、蛋白质聚集和炎症过程引起的损伤可能导致地图样萎缩和/或脉络膜新生血管形成和纤维化。近年来,细胞自噬在AMD病理中的作用逐步显现,东芬兰大学的Kai Kaarniranta教授等人汇总了相关资料,对选择性和分泌性自噬在AMD发病中玻璃膜疣的发生、衰老相关分泌表型和上皮-间充质转化(EMT)中的作用进行了详细阐述,有益于加深眼科医师对AMD发病机制的理解。
AMD病因探索
AMD病因由遗传因素和环境风险因素组成,包括衰老、吸烟、动脉硬化、肥胖、高血压、高胆固醇血症和富含脂肪的饮食。溶酶体清除中断和废物积累增加是导致AMD发生的主要细胞因素,自噬和泛素-蛋白酶体系统(UPS)这两种主要机制负责管理细胞中的废物清除。在很长一段时间里,学者们独立看待这些系统,但最新研究表明,导致AMD进展的清除缺陷应该在自噬和UPS相互作用的背景下考虑。
除了自噬参与导致AMD的细胞废物清除失调以外,其他一些因素,例如氧化应激、炎症和RPE死亡,也将自噬缺陷与AMD联系起来。活性氧(ROS)作为信号分子参与营养饥饿诱导的自噬,细胞抗氧化反应又由自噬通过SQSTM1/p62(螯合体1)-KEAP1-NFE2L2/Nrf2途径调节,SQSTM1在连接自噬和UPS方面发挥着特殊作用。另外,自噬也通过DNA损伤反应机制与抗氧化防御建立联系。
AMD中RPE变性的发生机制
RPE变性是AMD的特征之一,静止的RPE细胞形成单个细胞层,位于前部光感受器(视杆和视锥)与Bruch膜和绒毛膜毛细血管之间(图1)。RPE细胞的中心稳态功能之一是吞噬来自光感受器外节的视觉周期的富含脂质的终产物。这一过程可以解释为微管相关蛋白1轻链3(MAP1LC3/LC3)的相关吞噬作用(LAP,图2)。LAP的减少和线粒体衍生的ROS的过量被认为是触发AMD中RPE变性的关键因素。慢性氧化应激使蛋白质容易受到损伤,并导致异常蛋白质的有害积累,这对于RPE细胞等一部分分裂细胞的正常稳态尤为重要。蛋白稳定紊乱的一个明显临床迹象是溶酶体脂褐素在RPE中的积聚以及RPE和绒毛膜毛细血管之间的细胞外核膜沉积(图1、图2)。蛋白酶体和溶酶体在防止蛋白质聚集和细胞清除方面发挥主要作用。目前,新出现的证据表明,自噬在AMD发生和进展的各个阶段发挥着重要作用,由于溶酶体酶活性对自噬流量至关重要,脂褐素成分可能最终降低自噬清除能力。此外,自噬能力的下降伴随着ROS产生的增加,可以引发RPE细胞炎症的激活。
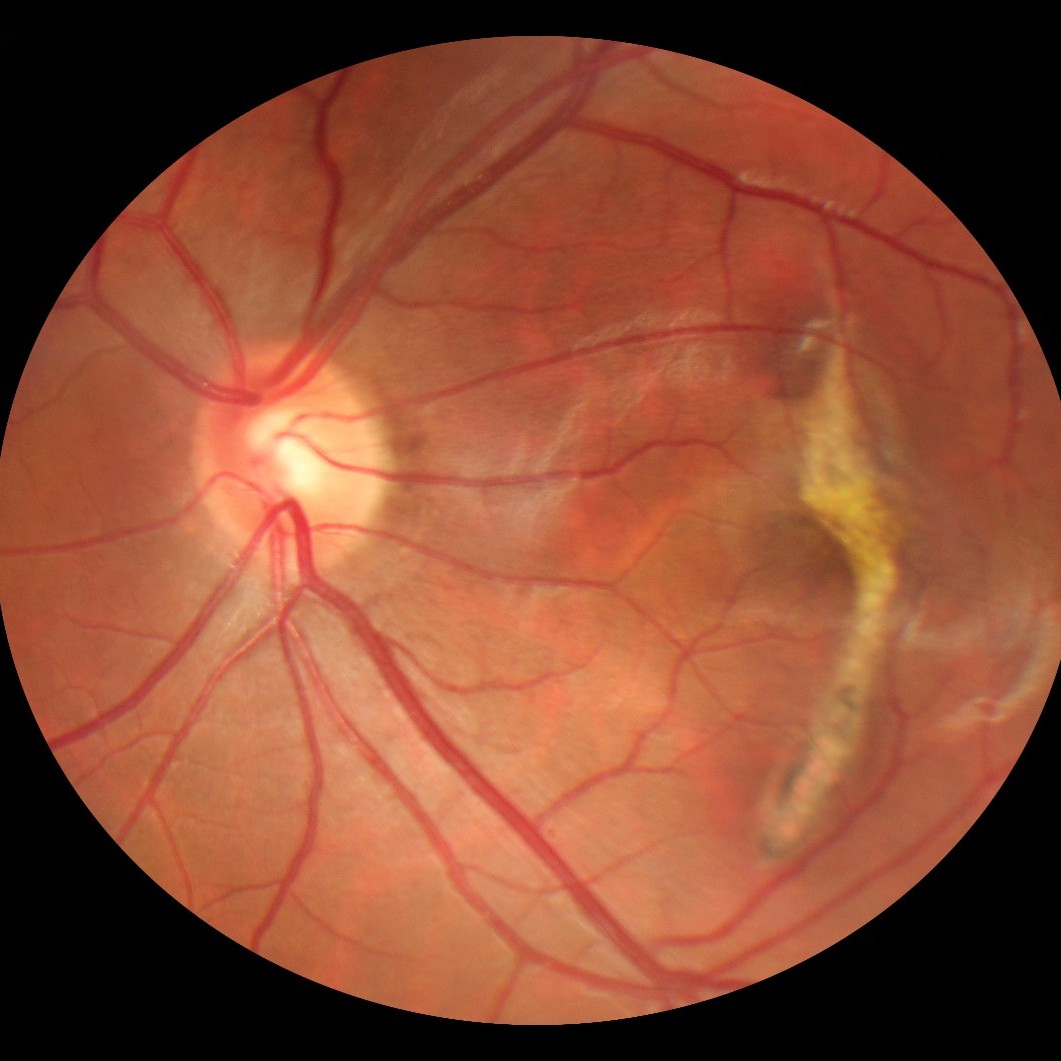
图1. (A)正常眼的彩色眼底照片(上图)和相干光断层扫描照片(下图);(B)伴有玻璃膜疣的干性AMD(白色箭头),以及(C)伴有出血、视网膜内、视网膜下和视网膜色素上皮下液的湿性AMD(红色箭头)。
图2. 自噬机制图解
AMD选择性自噬
自噬清除机制在AMD发病机制中的关键作用正在显现,简言之,雷帕霉素激酶的机制靶点(MTOR)、自噬相关基因(ATG)、unc-51样自噬激活激酶1(ULK1)、LC3和SQSTM1(以及其他受体蛋白)是调节选择性自噬的关键分子。由核心自噬基因ATG5和ATG7的缺失引起的自噬损伤与小鼠RPE细胞中的AMD样表型有关,这种表型表现为RPE增厚或营养不良、色素异常和氧化蛋白积累。氧化应激是RPE细胞诱导自噬的典型刺激,晚期AMD的自噬流量发生率低于疾病早期。慢性氧化应激似乎会降低RPE细胞中的溶酶体酶活性和自噬流量。
AMD线粒体自噬
RPE变性的确切原因以及AMD发病和进展机制尚不完全清楚,然而在AMD中,观察到线粒体功能障碍、ROS产生增加、线粒体DNA/mtDNA损伤以及蛋白质聚集和炎症增加。为避免大量ROS产生与线粒体进一步损伤和ROS释放的恶性循环,必须通过线粒体自噬将功能失调的线粒体从细胞中清除。PTEN诱导的激酶1(PINK1)、parkin RBR E3泛素蛋白连接酶(PRKN)和视神经磷酸酶(OPTN)是线粒体自噬途径的一些主要蛋白。一旦线粒体损伤发生,线粒体内膜去极化,PINK1输入线粒体,其被人早老素相关菱形样蛋白(PARL)降解受到抑制,导致其在线粒体外膜中的积累增加(图3),随后从胞质溶胶中募集PRKN。PINK1磷酸化泛素和PRKN在丝氨酸65的泛素样结构域,一旦被募集和激活,PRKN将泛素化许多靶标,并通过受体与LC3相互作用,通过其LC3相互作用区(LIR)基序刺激吞噬体局部形成。氧化应激显著增加了RPE细胞中参与线粒体自噬的关键分子(PINK1和PRKN),以分离受损的线粒体并通过自噬将其清除。
图3. 线粒体自噬机制图解
AMD内质网选择性自噬
慢性氧化应激、自噬受损、蛋白质聚集增加和RPE细胞变性与AMD病理中的内质网(ER)应激一致。为了维持蛋白稳定和细胞功能,ER激活一种名为未折叠蛋白反应(UPR)的适应性质量控制机制。许多移动到内质网、高尔基体、细胞膜或溶酶体的蛋白质在内质网膜上合成,这些蛋白质经过各种翻译后修饰获得最终的功能结构,在其氨基末端尾部具有ER前导序列的蛋白质通常通过高尔基体分泌到细胞外,无领导序列蛋白直接从细胞质中进行非常规分泌,常规和非常规分泌途径都与自噬相互作用。
有证据表明RPE细胞和AMD发展过程中存在自噬ER线粒体串扰,内质网-线粒体接触位点参与自噬体的形成,线粒体相关的内质网膜/MAM区室中的许多蛋白质是自噬小泡形成所必需。自噬途径还调节细胞因子的产生和分泌,特别是IL1B/IL-1β(白细胞介素-1β),反之亦然,包括IL1B在内的细胞因子诱导自噬并激活自噬体的形成。
AMD的自噬和衰老
自噬减少与细胞表型向衰老细胞转移有关,会促进组织稳态丧失。RPE中脂褐素的积累是AMD衰老的标志,衰老相关分泌表型(SASP)与ROS、选择性生长因子、炎症细胞因子、趋化因子和蛋白酶的释放有关(图4)。高迁移率组盒1(HMGB1)是一种衰老标志物,其在iPSC RPE中响应脂褐素组分处理而上调,线粒体损伤或内质网应激也上调HMGB1。AMD相关的细胞变化,如端粒功能障碍、DNA损伤和代谢紊乱,可能导致衰老细胞的持续积累。由于衰老细胞表现出与促炎分泌组一致的代谢功能和自噬活性紊乱,它们可能调节玻璃膜疣的发生和某些患者的晚期AMD发展。细胞衰老具有依赖于环境的有益或有害作用,但在慢性氧化应激和RPE变性过程中,衰老可能会发展为AMD。因此,去除衰老的RPE细胞或抑制其活性是AMD的潜在治疗选择。
图4. 变性RPE细胞中细胞质废物清除和分泌系统的相互作用
自噬在预防EMT中的作用
EMT是一个极化的上皮细胞将其结构和生化特性转变为更具运动性和适应性的间充质细胞的过程。自噬通过清除SNAI/snail和TWIST来预防EMT,从而诱导EMT的转录因子。RPE细胞很可能通过启动2型EMT逃离氧化损伤产生的应激微环境,这是避免细胞死亡的一种机制。然而,在AMD患者中,随着疾病的发展,细胞会由于损伤的慢性性质而死亡。RPE可能由于维持RPE细胞稳态所需的各种途径/过程的故障而开启EMT,对RPE细胞存活极其重要的一个过程是溶酶体介导的吞噬和自噬清除。
小结
Kai Kaarniranta教授最后总结道,最近的研究支持氧化应激增加、蛋白质聚集、细胞衰老、慢性炎症,以及一些患者的脉络膜新生血管和EMT表型在AMD发病机制中发挥作用,由于自噬在这些细胞过程的调节中起着核心作用,眼科医师应该更好地了解AMD背后视网膜生理和病理中的不同自噬机制,调节自噬的miRNA已被证明是AMD未来的治疗选择。未来眼科医师有必要应用精准医学,了解自噬调节因子的遗传、表观遗传学、代谢组学和蛋白质组学病因,以更好地预防或治疗AMD。
参考文献:Kai Kaarniranta, Janusz Blasiak, Paloma Liton,et al. Autophagy in age-related macular degeneration. AUTOPHAGY, 2023;19(2):388-400.
https://doi.org/10.1080/15548627.2022.2069437
2 comments



































































































































































 京公网安备 11010502033360号
京公网安备 11010502033360号
条评论
Linda Gareth
2015年3月6日, 下午2:51Donec ipsum diam, pretium maecenas mollis dapibus risus. Nullam tindun pulvinar at interdum eget, suscipit eget felis. Pellentesque est faucibus tincidunt risus id interdum primis orci cubilla gravida.